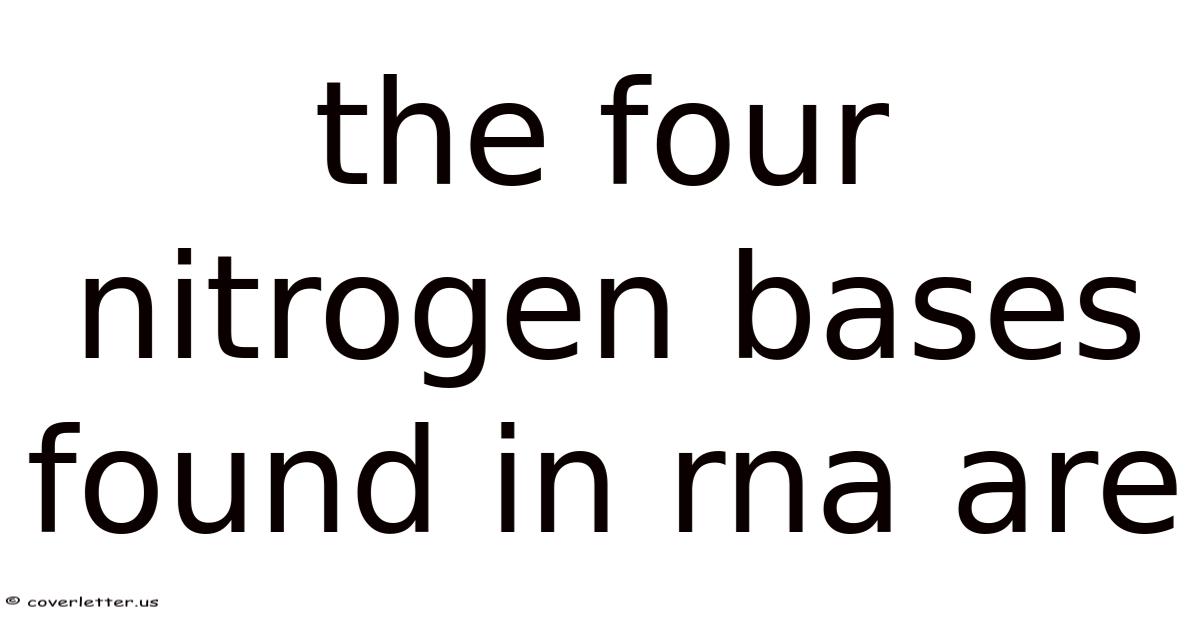
You've probably seen the letters A, U, C, and G scattered across biology textbooks, research papers, and those colorful DNA helix models. That said, maybe you memorized them for a test. Maybe you've stared at a sequencing readout and wondered what they actually do inside a living cell.
Here's the thing — most people can name the four bases. Far fewer can explain why RNA uses uracil instead of thymine, or how a single base swap can cascade into a genetic disease, or why the wobble position in tRNA matters for protein folding But it adds up..
Let's fix that.
What Are the Four Nitrogen Bases in RNA
RNA — ribonucleic acid — relies on four nitrogenous bases to store and transmit genetic information. They are:
- Adenine (A) — a purine, double-ring structure
- Guanine (G) — the other purine, also double-ringed
- Cytosine (C) — a pyrimidine, single-ring structure
- Uracil (U) — the other pyrimidine, single-ring, and the one that replaces thymine
That's the short list. But each base carries chemical quirks that shape everything from transcription fidelity to ribosome mechanics.
Purines vs pyrimidines — the structural split
Adenine and guanine are purines. Two fused rings — a six-membered ring fused to a five-membered ring. Which means bulky. Rigid. They take up more space in the helix.
Cytosine and uracil are pyrimidines. Smaller. One six-membered ring. More flexible.
This size difference isn't trivia. Too much gap. On top of that, try to cram two purines across from each other and the helix distorts. A pairs with U (two hydrogen bonds). Still, it's why base pairing follows Chargaff's rules — a purine always pairs with a pyrimidine. This leads to g pairs with C (three hydrogen bonds). The geometry only works that way. Two pyrimidines? The helix would buckle.
The uracil-thymine swap — why RNA ditched the methyl group
DNA uses thymine. A methyl group (-CH₃) at the 5-carbon position on thymine's ring. RNA uses uracil. Now, the only chemical difference? Uracil lacks it And that's really what it comes down to..
Seems minor. It's not.
That methyl group does two things. That said, first, it makes thymine more hydrophobic — better for DNA's long-term stability in the nucleus. In practice, second, it acts as a repair tag. Cytosine spontaneously deaminates into uracil at a measurable rate. So if DNA used uracil natively, the repair machinery couldn't distinguish "legitimate uracil" from "damaged cytosine. " By using thymine instead, any uracil that shows up in DNA is instantly flagged as an error No workaround needed..
RNA doesn't need that luxury. Still, it's transient. Most mRNA lasts minutes to hours. The cost of occasional mispairing is lower than the metabolic cost of maintaining a separate thymine synthesis pathway. Evolution took the shortcut Simple, but easy to overlook..
Why These Bases Matter — Beyond the Textbook Definition
You might ask: okay, four bases, two pairs, got it. Why does anyone spend a career studying this?
Because the consequences of those four letters ripple through every biological process Simple, but easy to overlook..
Information density and the genetic code
Four bases. Three positions per codon. 4³ = 64 possible codons. That's enough to encode 20 amino acids plus start/stop signals — with redundancy built in. So the degeneracy of the code (multiple codons for the same amino acid) isn't waste. It's a buffer against mutation. A single-base change in the third position often doesn't alter the amino acid. That's not luck — that's the wobble hypothesis in action, and it exists because of how these four bases pair.
Base modifications — the hidden layer
Here's what most intro courses skip: the four standard bases are just the starting canvas. Practically speaking, over 170 chemically modified nucleosides have been identified in RNA. Pseudouridine (Ψ), inosine (I), N⁶-methyladenosine (m⁶A), 5-methylcytosine (m⁵C) — the list goes on.
These modifications tune everything. Also, inosine at the wobble position expands decoding capacity. m⁶A acts as a "reader" tag for mRNA stability and translation efficiency. Pseudouridine in tRNA stabilizes codon-anticodon pairing. Plus, the four bases are the alphabet. Modifications are the punctuation, the accent marks, the formatting.
Ribozymes and the RNA world
RNA isn't just a messenger. It catalyzes reactions. The ribosome's peptidyl transferase center? Worth adding: pure RNA. Self-splicing introns? RNA. Which means rNase P? Think about it: rNA. These ribozymes fold into precise 3D shapes using the same four bases — plus magnesium ions and water molecules — to perform chemistry that proteins usually handle.
Not obvious, but once you see it — you'll see it everywhere.
The fact that four bases can build both information storage and catalytic machinery is a major pillar of the RNA world hypothesis. Life likely started with RNA doing both jobs before DNA and proteins took over the specialized roles But it adds up..
How It Works — From Transcription to Translation
Let's walk through the lifecycle of these bases in a eukaryotic cell. Not as a diagram — as a process with physical constraints.
Transcription — writing the message
RNA polymerase II slides along the template strand of DNA, reading 3'→5', synthesizing RNA 5'→3'. At each step, it selects the correct nucleoside triphosphate (ATP, UTP, CTP, GTP) based on Watson-Crick pairing with the DNA template.
A in DNA → U in RNA
T in DNA → A in RNA
G in DNA → C in RNA
C in DNA → G in RNA
The polymerase active site discriminates against deoxyribonucleotides (missing the 2'-OH) and against wrong bases. Fidelity is decent — error rate around 10⁻⁴ to 10⁻⁵ — but not perfect. That's why proofreading exists in some viral polymerases, and why cellular RNA surveillance pathways (nonsense-mediated decay, no-go decay) catch faulty transcripts later.
Splicing — editing the raw transcript
Pre-mRNA contains introns. The spliceosome — a massive ribonucleoprotein complex — recognizes splice sites via base pairing between snRNAs (U1, U2, U4, U5, U6) and the pre-mRNA. And the branch point adenosine attacks the 5' splice site in a transesterification reaction. That adenosine? It's one of the four bases, positioned by RNA-RNA pairing, acting as a nucleophile Most people skip this — try not to. Worth knowing..
No proteins catalyze the chemistry. The RNA does it Most people skip this — try not to..
Translation — reading the code
mRNA enters the ribosome. tRNAs deliver amino acids. Each tRNA has an anticodon loop — three bases that pair with the mRNA codon. The pairing follows Watson-Crick rules at positions 1 and 2. Here's the thing — position 3? Wobble.
Crick's wobble hypothesis (19
Wobble and beyond: how modifications fine‑tune the code
Crick’s wobble hypothesis (1966) proposed that the third base of a codon could pair flexibly with the first base of the anticodon. In practice, this flexibility is realized through non‑standard base pairing at the wobble position, which is often modified in tRNA. Inosine (I) is the classic example: it can pair with A, U, or C, allowing a single tRNA to recognize three synonymous codons. Other modifications expand the repertoire even further.
Pseudouridine (Ψ) replaces uridine in the anticodon loop and stabilizes the local helix through extra hydrogen‑bond donors and improved base‑stacking. Think about it: 5‑methyloxycarbonylmethyluridine (mcm⁵U) and its derivatives increase pairing strength with A or G, while thiolated bases such as 5‑thiouridine (s²U) bias pairing toward A and help discriminate against near‑cognate codons. On the flip side, n⁶‑threonylcarbamoyladenosine (t⁶A) improves Watson‑Crick pairing at the first anticodon position, reducing misreading. Collectively, these modifications act like a punctuation system: they clarify where a codon ends, point out certain pairings, and even introduce new “letters” to the genetic alphabet without changing the underlying DNA blueprint.
The impact of these tweaks reaches far beyond the ribosome. Modified nucleosides in mRNA—such as N⁶‑methyladenosine (m⁶A)—serve as signals for RNA stability, splicing, and translation efficiency. By recruiting specific reader proteins, m⁶A can accelerate decay of transcripts that carry premature stop codons, ensuring quality control. In bacterial mRNA, 2‑methylthio‑N⁶‑methyladenosine (ms²t⁶A) enhances ribosome processivity under stress conditions, while pseudouridines placed in coding regions can increase translational fidelity by stiffening the RNA backbone.
These layers of regulation illustrate that the genetic code is not a static script but a dynamic document. The four canonical bases provide the basic vocabulary, but chemical modifications add punctuation, accent marks, and formatting that shape how the message is read, repaired, and ultimately expressed Surprisingly effective..
From the RNA world to modern biotechnology
The same chemical versatility that allows a single tRNA to recognize multiple codons also underpins the catalytic prowess of ribozymes. The peptidyl‑transferase center of the ribosome, self‑splicing introns, and RNase P all rely on precise folding of RNA structures that are stabilized by metal ions, hydrogen bonds, and modified nucleotides. The fact that life could have emerged from a world where RNA performed both information storage and catalysis suggests that modifications were not later inventions but integral features from the start—nature
Building on this foundation, researchers have begun to harness modified nucleotides as programmable elements in synthetic biology. In parallel, orthogonal ribosome systems have been constructed in which the ribosome’s decoding pocket preferentially recognizes tRNAs modified with unique bases, allowing parallel translation of distinct mRNA repertoires within the same cell. By engineering tRNA synthetases that specifically charge tRNAs bearing non‑canonical anticodon–codon pairs, scientists can expand the genetic code to incorporate unnatural amino acids with novel side chains, thereby endowing proteins with unprecedented functions. Such modularity illustrates how subtle chemical alterations can create entirely new layers of regulation without rewriting the underlying DNA sequence Small thing, real impact..
The therapeutic potential of RNA modification has also come to the fore. In real terms, messenger RNAs used in vaccines and gene‑editing tools are deliberately enriched with pseudouridine and N¹‑methyl‑pseudouridine to dampen innate immune activation and boost translational efficiency. Practically speaking, by fine‑tuning the pattern of methylation on the 5′ cap and internal residues, developers can modulate the half‑life of the transcript, ensuring that the encoded protein is expressed at the optimal dosage and duration. Also worth noting, site‑specific incorporation of 5‑thiouridine or 2′‑O‑methyl groups can create “soft spots” that are selectively cleaved under disease‑associated conditions, offering a molecular switch that can be activated by endogenous enzymes.
People argue about this. Here's where I land on it.
From an evolutionary standpoint, the diversity of modifications provides a mechanistic explanation for the robustness observed in many organisms. Day to day, in fluctuating environments, the ability of a single tRNA to decode multiple codons through inosine or thiolated uridine can buffer translational errors caused by temperature shifts or nutrient scarcity. Likewise, the presence of methyl groups on the ribose or bases can protect RNA from degradation by nucleases, extending the functional lifespan of transcripts that encode stress‑response proteins. These adaptive advantages likely contributed to the conservation of modification enzymes across domains of life, reinforcing the view that they were present in the earliest replicators rather than emerging later as ancillary features Still holds up..
Looking ahead, the systematic mapping of the “epitranscriptomic” landscape promises to reveal further connections between chemical marks and cellular phenotypes. High‑throughput sequencing combined with bisulfite‑free chemistries now enables the detection of dozens of modifications directly from cellular RNA, opening avenues to correlate specific modification patterns with disease states, developmental stages, or environmental responses. As the field progresses, the distinction between a static genetic script and a dynamically edited document will become increasingly blurred, reinforcing the central message that the true power of genetic information lies not merely in its sequence but in the nuanced choreography of chemical modifications that dictate how, when, and with what fidelity that information is expressed Worth keeping that in mind..